
前面的三个例子,显示比较解剖学足以解答物种关系的问题,根据遗传证据得到的结论,只不过证实了解剖学家早已发现的事实。但是DNA也可以解决解剖学无法解答的问题:人、大猩猩与黑猩猩的关系。如图1所显示的,人类与两种黑猩猩,有1.6%的遗传差异,相同的基因达98.4%。大猩猩的差异较大,与人或黑猩猩的差异是2.3%。
另一方面,希伯利和阿基斯特发现黑猩猩与波诺波猿(过去叫倭黑猩猩)的DNA最相近,只有0.7%的差异。这也不令人感到意外。这两种猿看来非常相似,直到1929年才有解剖学家觉得该为它们分别取个名字。生活在前比属刚果中部的“黑猩猩”,是倭黑猩猩,因为一般来说它们体型较小、体格稍瘦、两腿较长。一般的黑猩猩,在非洲分布较广,主要在赤道以北。然而,对这两种黑猩猩的行为,有了比较详细的记录后,学者才恍然大悟,原来它们形态上并不起眼的差异,掩盖了生殖生物学上的重大差异。倭黑猩猩——现在叫波诺波猿——不像黑猩猩,倒像人,它们有很多种性交姿势,包括面对面式;两性都会主动挑逗对方,而非总是雄性主动;雌性并非只在“发情期”(排卵期)接纳雄性,几乎整个生殖周期都能性交;雄性之间、异性之间都能“结盟”,而不限于雄性。很明显地,那0.7%的遗传差异,已在性生理与行为上造成了重大的结果。在本章与下一章,我们会反复强调少数基因的重大后果。
让我们在这儿稍作逗留,仔细咀嚼这几个数字的意义:
他们的第二个发现,也不令人意外:长臂猿与人/猿的DNA,有5%的差异。这也证实了学界的共识:长臂猿是最特殊的猿;与我们关系比较密切的猿是大猿,像红毛猩猩、大猩猩、黑猩猩等。在大猿中,最近解剖学家已经开始认为红毛猩猩很早就自成一家,这与DNA证据也很吻合,它的DNA与其他大猿以及人的,有3.6%的差异。生物地理也支持非洲大猿与亚洲猿(长臂猿、红毛猩猩)很早就分家了。现在,长臂猿、杂毛猩猩只分布在东南亚,它们的祖先化石也只在东南亚出土;而大猩猩与两种黑猩猩,只分布在非洲,人类早期祖先的化石,也只在非洲出土。
在我们的族谱上,大猩猩必然在我们与黑猩猩分家前,就分离出去了。我们最亲近的亲戚,是黑猩猩,而非大猩猩。另一方面,黑猩猩最亲近的亲戚,是人,而不是大猩猩。传统分类学将所有大猿放在同一分类类目中(“猿科”),为人单独另立一个类目(“人科”),好像人与猿之间有一不可逾越的自然鸿沟,对我们自居于“万物之灵”的“人本位”偏见,有推波助澜之功。现在呢?未来的分类学家也许可以用黑猩猩的眼光来处理高等灵长类的分类问题:把它们大致分为两群,一群包括三种黑猩猩(人加上另外两种黑猩猩),另一群包括其他的猿(长臂猿与红毛猩猩与大猩猩)。两群之间并没有明显的差异,三种黑猩猩那群只不过有点儿高明而已。传统分类学将人与猿分别开来,不符合事实。
正如解剖学家预测过的,他们发现的最大遗传距离,出现在人类或任何猿类与猴子之间,也就是说,人/猿或猿/猴的杂种DNA,“融解”的温度最低。这只不过是在大家都已经同意的看法上加个数字而已——自从科学界知道猿类存在之后,就认为猿比猴更接近人类。那个数字是7%:猴的DNA与人/猿的DNA,93%是相同的。
人与黑猩猩的遗传距离(1.6%),是两种黑猩猩的差异(0.7%)的两倍多,比两种长臂猿的差异小(2.2%)。红眼与白眼缘鹃是两种非常相似的北美鸟儿,也有2.9%的差异。我们的基因组中,98.4%的基因,都与黑猩猩的一样。举例来说,我们的主要血红素是红血球中携带氧气的分子,由287个氨基酸组成,与黑猩猩的一模一样。在许多方面,我们不过是第三种黑猩猩,对其他两种黑猩猩有利的,对我们也有利。我们看来与它们不同,是因为我们以直立姿态行走,脑子比较大,能说话,体毛稀少,有奇异的性生活,这些特征必然是我们基因组中那1.6%基因控制的。
图1 追溯每组现代高等灵长类至连接他们的黑点。左边的数字给出了那些现代灵长类的DNA差异百分比,而右边的数字给出了自他们最后拥有共同祖先的年代。例如,黑猩猩和波诺波猿的DNA差异是0.7%,他们在300万年前分化;我们与他们各自的DNA差异是1.6%,大约在700万年前与他们共同的祖先分化;大猩猩与三种黑猩猩的DNA差异是2.3%,大约在1000万年前与他们共同的祖先分化开来。
要是物种间的遗传距离以固定速率累积,遗传距离就可当作运转正常的时钟。将遗传距离转换算成绝对时间(两个物种从最后一个共祖分化出来,到现在所经过的时间),我们得找到一对物种,一方面它们的遗传距离可以测量,另一方面它们有年代确定的化石可供参考。事实上,高等灵长类有两组独立的换算数据。一方面,根据化石记录,2000万年到3000万年前与猴分家,两者的DNA差异达7.3%。另一方面,红毛猩猩1200万年到1000万年前与其他大猿分化,DNA差异达3.6%。比较这两组数据可以发现:演化时间增加了一倍(从1200万年到2500万年),差异就增加一倍(从3.6到7.3)。因此,高等灵长类的DNA时钟,运转得相当稳定。
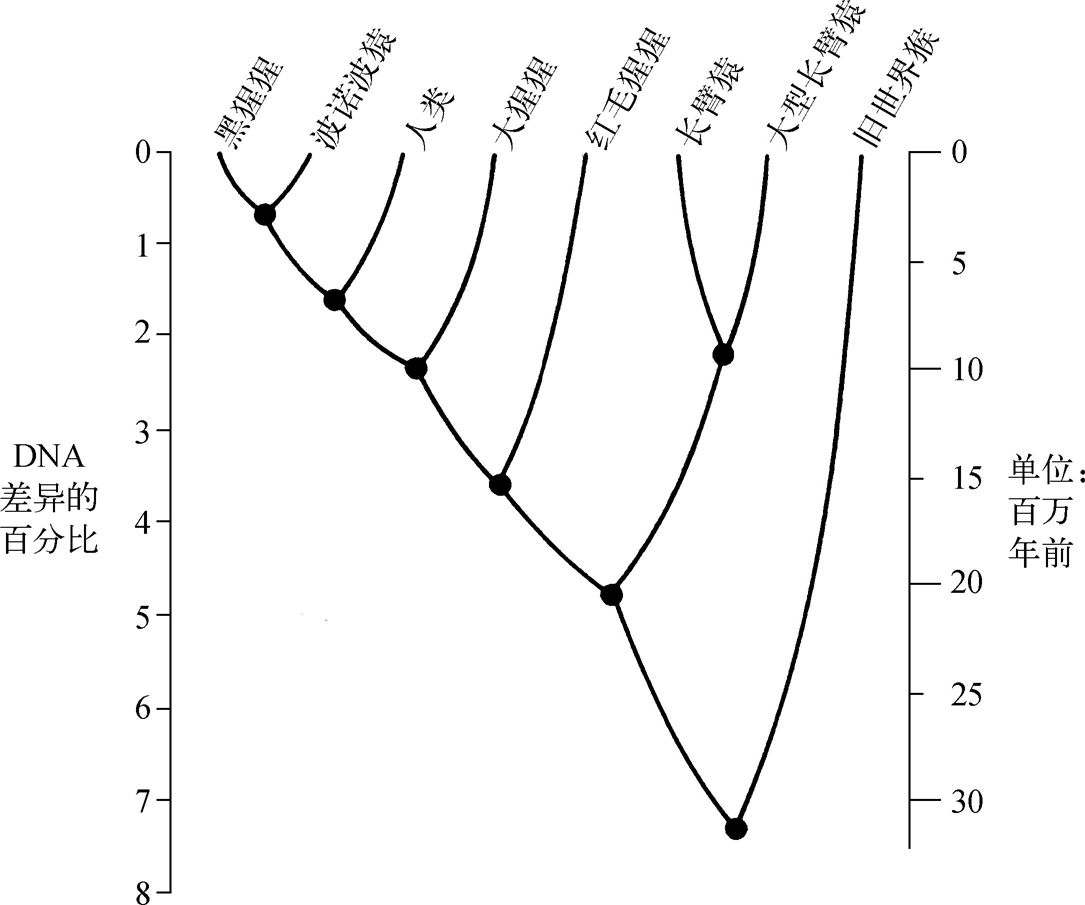
于是,希伯利和阿基斯特以那些换算尺度,估算我们的演化史。由于我们与黑猩猩的遗传距离(1.6%),是红毛猩猩与黑猩猩的一半(3.6%),因此我们与黑猩猩分别演化的时间,只是红毛猩猩与黑猩猩的一半(1200万年到1600万年的一半)。换言之,人与另外两种黑猩猩,约600万年到800万年前分别走上不同的演化道路。同样地,大猩猩与黑猩猩分化的时间,以及黑猩猩与波诺波猿分化的时间,我们都可以算出来,分别是900万年前与300万年前。你知道吗?我上大一时(1954年)的体质人类学教科书,说人与猿1500万年到1300万年前就分家了。因此,DNA时钟支持一个引起争议的结论,其他好几种分子时钟(例如蛋白质氨基酸序列、粒腺体DNA等)也得到同样的结论。每一种时钟都指出:人类最近才与黑猩猩分化,是个年轻的物种,比古生物学家过去所推测的年轻得多。
高等灵长类的族谱
这些结果对于我们在动物界的地位,有什么意义?生物学家将现生生物分门别类,各从其类。分类系统是个层级体系:亚种、种、属、科、目、纲、门、界,一层比一层“笼统”。《大英百科全书》与我书架上所有生物学教科书,都说人与猿属于同一目(灵长目),同一超科(人超科),但不同科。人属于人科,猿属于猿科。希伯利和阿基斯特的研究会不会改变这个分类,视学者的分类哲学而定。传统的分类学家,将不同物种归入同一个较高层次的类目,使用的方法不免主观,因为得从诸多相似相异之处分别主次,找出重要的差异(相似),忽视其余。这样的分类学家,会因为人类拥有独特的功能特征(大脑与直立姿态),而为人类单独立一类目,希伯利和阿基斯特的结果不会改变他们的结论。
当年达尔文在讨论“人类歧异”这个爆炸性议题之前,花了近10年研究藤壶(一种水栖节肢动物)的歧异。同样地,希伯利和阿基斯特也花了10年,以DNA时钟厘清鸟类的关系。1984年,他们第一篇以DNA时钟讨论人类起源的论文发表了。此后他们出版了一系列论文,更精炼了当初的结论。他们取得的DNA,包括人类的,以及所有人类的近亲——红毛猩猩、大猩猩、黑猩猩、波诺波猿,还有两种长臂猿、7种旧世界猴。图1概括了此结果。
然而,另一个分类学派,叫做分支系谱学派,则认为生物分类应该遵循客观一致的程序,遗传距离或分化时间是惟一标准。所有分类学家都同意:红眼缘鹃与白眼缘鹃都是“缘鹃属”的鸟种:所有长臂猿都属于“长臂猿属”。然而这两个属的成员,彼此的遗传距离,却大于人与黑猩猩的,而且早就分化了。因此,人类就不可能独立成科,甚至不应独立成属,而应与另外两种黑猩猩归入同一属。可是根据动物命名国际公约,人属(Homo)这个属名比较早问世,因此人属这一属应有3个物种,除了人,还有黑猩猩、波诺波猿。由于大猩猩只不过更独特一些,因此它几乎可以算成人属中第四个物种。
依我之见,尘埃落定之后,那位遗传学家的意见最接近真相。DNA时钟的原理,无可挑剔;希伯利和阿基斯特使用的方法是最先进的;他们测量过18000对“杂种”DNA,得到的遗传距离,呈现了内部的一致性,证明他们的结果是恰当的。
即使分支系谱学派的分类学家,也免不了“人本位”偏见,他们对“人类与非洲大猿共处一个类目”的结果,想必也觉得是良药苦口、难以下咽。不过,一旦黑猩猩学会了分支系谱派分类学,或外太空来了个分类学家,一定会毫不犹豫地接受新的分类。
“他们的结果是‘上帝的启示’,你最好相信。”(一位遗传学家)
人与黑猩猩有哪些基因不同?回答这个问题之前,我们得了解DNA(遗传物质)是做什么的。
“有意思,可是他们的结论必须通过其他方法的验证,我们才会相信。”(一位演化生物学家)
我们的DNA大部分都没有功能,有的话,科学家还没有发现。它们也许只是“分子垃圾”:或者一个基因复制了好几份,其中一份继续发挥功能,其他的几份就无所事事充场面;或者是丧失了功能的基因。总之,它们没用,可是又无害,所以没被淘汰。我们的DNA有功能的部分,主要与合成蛋白质有关:蛋白质是氨基酸组成的长链分子。有些蛋白质大部分身体结构都有,例如角蛋白(皮肤、毛发、指/趾甲)与胶原(结缔组织中的成分)。不过另有一群蛋白质,负责合成或分解身体里的分子,我们通称为瓷。DNA分子上的核苷酸顺序,是制造蛋白质的指令。因为根据DNA分子上的核苷酸,可以调动不同的氨基酸组成需要的蛋白质。DNA分子上有功能的部分,有些负责合成蛋白质,另外的就负责调节合成蛋白质的工作。
“他们的方法倒没问题。可是有什么人会想做那样沉闷的研究?鸟类的分类学?!难以想象!”(一位分子生物学家)
在我们的生物特征中,最容易以遗传机制来理解的,就是个别的蛋白质,以及制造那些蛋白质的基因。例如血红蛋白(前面已经提过,是我们血液中携氧气的蛋白质)包括两条氨基酸链,每一条都由一小段DNA(一个基因)负责制造。那两个基因除了制造血红蛋白中的氨基酸链之外,并没有其他观察得到的功能。而血红蛋白只有红血球才有。反过来说,血红蛋白的结构完全是由那两个基因决定的。不管你吃什么,有多少运动量,最多也是影响身体制造血红素的数量,不会影响血红蛋白的结构。
“我对他们那套玩意已没有什么耐心。不管他们写什么,我都懒得再理会。”(一位解剖学家)
那是最单纯的情况。但是有些基因会影响许多观察得到的特征。例如“泰萨二氏病”(Tay-Sachs disease)是一种致命的遗传疾病,有许多解剖与行为的症状:特别嗜睡、姿势僵硬、皮肤泛黄、头骨畸形发育等。科学家发现:所有那些症状都是因为一个由“泰萨二氏基因”决定的酶,发生了变化才产生的。至于那一个酶怎么捅下那么大的娄子,科学家还没搞清楚是怎么回事。因为那个酶在许多身体组织中都有,广泛地参与了许多细胞成分的分解,难怪一旦改变了,会产生那么多症状,最后让病人送命。不过有些身体性状是由许多基因共同控制,例如身高,环境因素也扮演了一些角色,例如发育阶段的营养状况。
虽然希伯利和阿基斯特的成就,已是鸟类分类学的里程碑,但他们起先在学界引发的不是赞辞,而是批评。因为当时有几位科学家有足够的背景知识,能做中肯评论的人少之又少。我就听过科学界的同僚发表过这样的高论:
负责制造已知蛋白质的基因,科学家已经发现了许多,对它们的功能也很清楚。不过对涉及复杂性状、特征(例如大部分行为)的基因,却所知不多。像艺术、语言或暴力等人类特色,绝对不可能只由一个基因负责。人与人之间的行为差异,明显地受环境的强烈影响,基因扮演的角色一直受到争议。不过,黑猩猩与人的行为差异,倒可能涉及遗传差异,虽然现在还无法具体指出哪些基因牵涉在内。举例来说,人类能说话,黑猩猩就不能,控制声带构造与大脑神经网络的基因,必然是关键。曾有一对心理学家,收养了一头黑猩猩婴儿,与自己的新生儿一起抚养。他们受到一视同仁的待遇,吃、喝、穿、住都一样,以及同样的“教育”。结果,黑猩猩婴儿长大了,不会说话,也不能像人一样,直立走路。但是一个人长大后说什么语言,英语还是汉语,就不是基因决定的,孩子发育期间的语言环境,是惟一的决定因素。在美国出生的华人,能说一口流利的美语,已经不是新闻,不必再举例了。
在20世纪70年代,分子生物学家与分类学家大多数都对彼此的研究不感兴趣,只有少数例外。耶鲁大学的希伯利(Charles Sibley)是其中之一,他是鸟类学教授,兼任耶鲁大学皮博帝自然历史博物馆馆长。鸟类的分类学不容易研究,因为鸟类的身体是为飞行设计的,而设计一只鸟也不过就那么几种花样,即使大自然也出不了新招。所以在类似环境中生活的鸟,往往形态非常相似。例如在半空捕食昆虫的鸟,即使关系疏远,比较解剖学的差异也不大。美洲秃鹰与旧世界秃鹰,形态与行为都很像(可是学者好不容易才搞清楚美洲秃鹰与鹤鸟比较亲近,而旧世界秃鹰与老鹰比较亲近),是由相同的生活形态造成的。希伯利与阿基斯特(Jon Ahlquist)由于深切体会到传统分类方法的短处,在1973年开始采用分子时钟技术。当年他们应用分子生物学方法,解决分类学问题,就研究规模而言是空前的。他们在1980年开始发表研究结果,他们用分子时钟技术累计测量过1700种鸟,占世界鸟类的1/5。
有了这个基本认识之后,让我们再回头讨论人类与黑猩猩那1.6%的遗传差异。我们知道制造主要血红素的基因并没有改变,其他的基因有一些有很小的变化。人类与黑猩猩都有的9种蛋白质,共由1271个氨基酸组成,其中只有5个彼此不同:一个出现在肌球蛋白,一个在次要血红素的丁链上,3个在一种酶上(carbonic anhydrase,碳酸酐酶)。但是,第二章到第七章我要讨论一些人与黑猩猩的重大功能差异,如脑容量、骨盆、声带、生殖器的构造、体毛、女性月经周期,以及停经等。它们由哪些基因负责,我们还没有头绪。上面提到的5个氨基酸差异,不可能造成那么重大的后果。现在我们可以肯定的是:我们的DNA盘踞着大量“垃圾”;我们与黑猩猩那1.6%的遗传差异中,也有垃圾:我们与黑猩猩的重大功能差异,是那1.6%中的一小部分造成的。
分子生物学家在1970年左右解决了前两个问题。他们发现最适合的分子是脱氧核糖核酸(DNA);沃森与克里克1953年证明这个分子的构造是双螺旋,为遗传学研究开辟了新天地,也使DNA成为家喻户晓的分子。每个DNA分子包含两条互补的长链,每条链都由4种更小的分子单位组成,这4种分子在链上的顺序,蕴涵着从父母亲传递下来的所有遗传信息。科学家发展了“DNA杂交”技术,可以迅速测量DNA的变化。先将两个不同物种的DNA分子分离(“融解”)开来,就是使每个DNA分子的两条长链解开,分别独立。再让那些单链“DNA杂交”成为双链的DNA。然后加热,使“杂种”DNA再度分离开来。一般而言,需要的温度越高,就表示这两种DNA的结合程度越好,也就是彼此的差异越小。两个物种亲缘关系越近,DNA的差异越小。以“融解”一个物种的DNA的温度为基准,融解“杂种”DNA所需的温度,比基准度每低一度,表示两个物种的DNA有大约1%的差异。
总之,我们的DNA中,只有极小比例的基因在演化过程中改变了,其中一些对我们的身体产生了重大功能影响。并不是所有的基因变化都会产生同样的后果,因为大部分氨基酸,至少可以由两种核苷酸顺序决定。因此,DNA上的核苷酸变化——“突变”——如果不影响对应的氨基酸,就等于没有变化,学者叫做“沉默的”突变。即使突变不沉默,真的造成对应氨基酸的变化,蛋白质的功能会不会因此改变,仍是个问题。有的氨基酸,化学性质相似,互换后不影响蛋白质的功能。若不是处于“敏感”地位的氨基酸,即使被性质差异很大的氨基酸替换了,也不会有了不得的后果。
举例来说,假定根据化石证据我们推断狮与虎(都是猫科)在500万年前分化。再假定它们的同一种分子,结构上有1%的差异。要是我们任选两个演化关系并不清楚的物种,发现它们的分子结构有3%的差异。那么分子时钟就会告诉我们,它们在1500万年前就分化了。用这个方法纸上谈兵,听起来很精彩,能不能通过实例的考验呢?生物学家为此忙了好一阵。应用分子时钟之前,要完成四件事:科学家必须找到最适合的分子;找到测量分子结构变化的方法,必须简单而迅速;证明分子时钟运行稳定(也就是,分子结构在相关物种体内以同一速率演化);测量分子演化率。
但是,蛋白质上决定功能的部分,若有一个氨基酸被性质大不相同的另一个氨基酸替换了,就可能造成明显的后果。例如镰刀形红血球贫血症,是可能致命的基因疾病,病人的血红素不正常,只是因为血红素287个氨基酸中,有一个被性质大不相同的另一个氨基酸替换了。原来病人的DNA上,对应那个氨基酸的3个核苷酸,有一个发生了变化(点突变)。原来的氨基酸带负电,取代它的不带电,血红素分子的电荷因此改变了,生化性质也就随之变了。
这些关于我们起源的问题,解决的方法来自一个始料未及的领域:运用分子生物学解决鸟类的分类问题。大约40年前,分子生物学家发现;动植物体内的化学分子可以当作“时钟”,用来测量遗传距离,以及两个物种在演化史上的分化时间。其中的逻辑是这样的:假定有一类分子,所有生物体内都有,它们在每一个物种中都有特定构造,而那些构造是由遗传密码决定的。再假定分子的构造会因为遗传突变而逐渐变化,而在所有物种中,变化率都一样。源自同一共祖的两个物种,体内的那个分子,起先构造应该完全一样,因为都是从共祖那里遗传来的。但是这两个物种分别演化以后,基因组中的遗传突变就各自独立累积,使那个分子的结构逐渐变化。因此,这两个物种的那个分子,会逐渐出现结构差异。要是我们能够算出平均每100万年会发生多少结构变化,任何两个有亲缘关系的物种,在分子结构上的差异,就可以当作一个“时钟”,来计算这两个物种已经和共祖分化多久了。
虽然我们不知道哪些基因或DNA上哪些核苷酸段落,造成区别人与黑猩猩的差异,可是我们有许多例子演示了“一两个或几个基因突变造成的巨大冲击”,“泰萨二氏基因”突变后造成许多重大而明显的症状,是其中之一。一个基因突变,改变了一个酶,居然产生了多方面的后果,进而夺人性命。一个基因突变,可以使同一物种的成员分别开来(病人/正常人)。有密切亲缘关系的物种呢?最好的例子是丽鱼。在东非的维多利亚湖,大约有200多种丽鱼,这种鱼是淡水鱼,在水族馆常见。根据学者研究,维多利亚湖的丽鱼,都是从20万年前的祖先种演化出来的。那200多种丽鱼,按照栖境来分类的话,可以找到老虎与乳牛的差别:以藻类维生的,捕食其他鱼的,咬碎蜗牛的壳吃里面的肉的,吃浮游生物的,捕食昆虫的,还有能将其他鱼的鳞片一点一点咬掉的,甚至专门捕食孵卵母鱼身旁胚胎鱼的。然而它们的平均遗传差异,只有0.4%。也就是说,使虎型摄食习惯转变成牛型习惯所需要的基因突变,甚至比把黑猩猩变成人的还要少。
到如今,解剖学家仍在辩论第一个问题:我们族谱上的细节。不过,不管解剖学家心中的人类演化族谱是什么模样,光凭解剖学研究,无法解答第二、第三个问题,也就是人和猿的分化时间与遗传距离。不过,也许化石记录在原则上可能解决族谱与分化时间的问题(遗传距离免谈)。要是我们有丰富的化石,举例来说,要是我们可以用一系列已断定了年代的化石,代表人类演化的各个阶段;另外还有黑猩猩的,也有大猩猩的。那么,也许我们会发现人与黑猩猩的化石系列,在1000万年前交会了,也就是找到人与黑猩猩的共祖了。而那一位共祖的演化系列,在1200万年前又与大猩猩的演化系列交会。可惜我们没有那么多的化石,尤其是现生猿类祖先的化石极为稀少。500万年前到1400万年前之间,是人类与非洲大猿演化的关键期,那个时段的猿类化石,尤其稀缺。
新的遗传证据,除了涉及分类学的技术问题之外,还有更深远的意义吗?也许最重要的,是让我们重新思考人与猿在宇宙中的地位。有名为万物之母,名字不只是技术细节的代号,还反映并创造态度。(你若不信,今晚请试试用“亲爱的”或“死猪”称呼你的另一半,记得要用同样的表情和语气。)新证据并不规定我们应该如何思考人与猿。但是,新证据可能会影响我们思考的方向,达尔文的《物种起源》就发挥了这样的影响,我们可能还要花上许多年才能把态度调整过来。在可能受到影响并产生争议的领域中,我只讨论一个例子:我们利用猿的方式。
在主流派中,大部分生物学家过去认为大猩猩与黑猩猩相似程度较高,也就是说,它们与人类都不大相似。换言之,大猩猩与黑猩猩还没来得及分化开来,我们就已经与它们分道扬镳了。这个结论反映了常识观点:大猩猩与黑猩猩可以归为一类,它们叫做猿,而我们则是人。不过,另外还有一种可能,那就是:我们看来与其他的猿不一样,是因为我们走上了一条独特的演化道路,我们的祖先自从与其他的猿分化之后,就在几个重要的方面发生了变化,例如行进的体态与大脑的容量。而大猩猩与黑猩猩却没有发生过什么重大变化,与当年的人猿共祖,模样没什么大差别。如果那是实情,人类可能与大猩猩最亲近,或与黑猩猩最接近,或者三者亲近的程度一样,彼此的遗传距离一样。
现在,我们认为动物(包括猿)与人之间,有根本的分别,我们的伦理规范与行为以这个分别为准则。举例来说(我在本章开宗明义,已经提过),我们可以将猩猩关进动物园的笼子里,公开展览,可是不能那么对待人。我常常想,要是动物园黑猩猩的笼子边上,分类名牌上注明的是“人属”的话,观众会有什么感受。然而,要不是公众在动物园里油然生出对猩猩的同情,保护野生猩猩的募款活动,也许不会得到热烈的社会响应。
起先,我们很自然地假定:比较解剖学早已解答了上面第一个问题。我们与大猩猩、黑猩猩特别相像,可是又与它们有明显的差别,例如我们的脑容量比较大、我们以直立的姿态行走、我们的体毛极少,其他还有许多差异,不过不是那么一目了然。然而,要是我们观察得仔细一点,这些解剖学事实并不能一劳永逸地解答我们的问题。同样的解剖特征,学者赋予不同的意义;而不同的学者,强调的解剖特征也可能不同。于是有的学者认为我们与亚洲猿(红毛猩猩)的关系最近,而大猩猩、黑猩猩是猿类中早就分化出的苗裔(少数派);有的学者认为我们与非洲猿(大猩猩、黑猩猩及波诺波猿)比较亲近,红毛猩猩是猿类族谱上最早分化出的猿(主流派)。
我也提到过,我们以黑猩猩做医学实验,既没征求它们同意,实验有时还有致命风险,可是没有人认为这有什么问题。换了人,就不可以。而以黑猩猩做实验,正是因为它们与人在遗传上非常相似。它们会感染许多人类的疾病,它们的身体对病媒的反应也与人相似。因此,以黑猩猩做实验,比用其他动物更能得到有用的资料,增进人类的医疗福利。
三、我们与关系最近的猿有多大的遗传差异?
这一伦理抉择,引发的问题更棘手,把猩猩关进动物笼子里,相形之下,还不算什么。因为我们将成千上万的人类罪犯关进牢里,已是例行公事。可是动物医学实验却没有“人类版本”。我们不是不知道:在人类身上做致命的实验,能得到更有价值的资料,用黑猩猩怎么也得不到。然而纳粹集中营医师以人类做实验,却受到各界的批评,认为是纳粹暴行中最可怕的罪行。为什么黑猩猩就可以?
二、不论哪一种现生猿与人类的关系最密切,人与那一种猿的最后一个共祖,在多久以前仍然活在地球上?
要是所有的生物,从细菌到人,可以排成一长列,我们必须决定在哪儿“杀”变成“谋杀”,“进食”变成“自相残杀”。大多数人将这条界线划在人与所有其他生物之间。不过,有不少人吃素,任何动物都不吃,可是吃植物。还有一小撮声音越来越大的人(属于为动物争权利的阵营)反对动物实验,或者说,反对以某几种动物做实验。主张动物权的人士,对猫呀狗呀或者灵长类,特别来劲,不怎么过问老鼠,而且一般而言,不为昆虫和细菌发言。
一、族谱:人类、现生猿类与化石“猿人”(类似猿的人:指人类的祖先)之间的亲缘关系是怎么样的?举例来说,要是我问你:“现生猿中,哪一种与人的关系最密切?”你怎么回答?
要是我们的伦理规范,在人与所有其他生物之间划下一条毫无道理的界线,那套规范摆明了就是私心作祟的产物,丝毫不含高尚的情操。要是我们的伦理规范,强调的是智慧、社会关系与感觉痛苦的能力,就很难在所有的人与所有其他生物之间,划下一条界线。那样的话,以不同的物种做实验,就要受不同的伦理规范监督。与我们有亲近遗传关系的物种,能不能享有特别权利呢?也许为它们大声疾呼的人士,也是出于私心,只不过戴上了新的面具。可是基于我刚刚提到的那些考量(智慧、社会关系与感觉痛苦的能力等),我们可以提出客观的主张,让大猩猩、黑猩猩享有“最惠物种”待遇。目前医学研究使用的动物中,要是有任何一种,我们可以合理地为它们申请保护令,不让它们再接受医学实验,那一定就是黑猩猩了。
在灵长类中,我们也很容易找到人类的地位,我们很明显地与猿比较相似,与猴的差异比较大。我只要指出一个最明显的特征就够了:猴子有尾巴而猿没有,我们也没有。猿这群灵长类中,长臂猿是最特殊的,它们体型小、手臂长,又叫小猿;其他的猿,如红毛猩猩、大猩猩、黑猩猩及波诺波猿都是大猿,它们彼此有很近的亲缘关系,与长臂猿比较疏远。但是想要进一步厘清我们与猿的关系,却非常困难,这也是始料未及的。这个问题还在科学界引发了热烈的辩论,论战围绕着三个议题:
动物实验造成的伦理困境,被黑猩猩搞得更严重,因为黑猩猩是濒临绝种的动物,医学研究不仅牺牲个体,还威胁了群体(物种)的命运。医学研究并不是威胁野地黑猩猩族群的惟一因素;栖境的破坏与动物园的需求,才是主要的威胁。但是研究的需求已构成相当程度的威胁,这就够医学界反省的了。还有其他的考量,使我们的伦理困境更显得阴郁:活捉一头黑猩猩,再将它送进医学实验室,整个过程中,平均起来会死好几头野生黑猩猩(往往是跟着母亲的幼仔);保护野地黑猩猩族群,生物医学科学家没出过什么力,虽然那么做怎么说都符合自己的利益;用来作研究的黑猩猩,往往没有受到人道的待遇。我第一次遇见供医学研究的黑猩猩,是在美国国家卫生院,它被注射了慢性的致命病毒,单独关在室内的笼子里好几年,连个玩具都没有,一直到死。
我们在动物界的地位,大体而言,学者早已确定,不知有多少世纪了。我们无疑是哺乳类,哺乳类的特征包括身体有毛发覆盖、哺乳等。在哺乳类中,我们属于灵长类,灵长类包括猴与猿等大类。我们与灵长类其他成员有许多共同的特征,例如指(趾)甲扁平而没有形成爪、有抓握能力的前爪、大拇指可以与其他手指对立、下垂的阴茎(而不是贴近腹部),这些特征大部分其他哺乳类都没有。早在公元2世纪,集西方古代医学大成的盖伦[1]从动物解剖经验中,已经正确地推定了人在动物界的地位,他发现猴子“无论在内脏、肌肉、动脉、静脉、神经,还是在骨骼形态上,都与人非常相似”。
人工繁殖黑猩猩供研究用,可以逃避危害野地黑猩猩族群的指控,可是仍然无法突破困境。19世纪美国禁止从非洲(或海外)输入黑奴,于是就有人贩售美国黑人的子女当奴隶,那可以接受吗?为什么可以用Homo troglodytes(黑猩猩)做实验,就不能用Homo sapiens(人)?反过来说,要是一个孩子得了有致命风险的病,我们正在以黑猩猩研究那种病,我们如何向孩子的父母解释:他们孩子的命,比不上黑猩猩重要?到头来,我们大众得做这些痛苦的抉择,而不是科学家。惟一可以肯定的是:我们看待人与猿的观点是关键。
也许你的回答是:猿是“动物”嘛,而人是人,这还有什么好讨论的。这是一条伦理准则,只有人类适用,不可引申到“动物”身上,不管那是一种什么样的动物,不管它与我们在遗传上多相像,不管它发展社会关系的能力多高强,不管它会不会感受痛苦。那个答案当然是武断的,可是至少它内部有一贯的逻辑,因此不可随便否定。不过这么一来,追溯我们的自然根源,便没有什么伦理意义。可是这样的探讨,仍能满足我们追求知识的好奇心,毕竟“我们从哪里来”这个问题是这么产生的。每个人类社群都对自己的起源有深切的好奇心,每个人类社群都有自己的创世故事。“三种黑猩猩”的故事,是我们这个时代的创世故事。
最后,改变我们对待猿的态度,也许是决定野地黑猩猩命运的关键。现在,它们的生命面临严酷的考验,特别是它们在非洲与亚洲的雨林栖境正遭到空前的破坏,它们被合法、非法地捕捉与猎杀。要是目前的趋势持续下去,不出20年,高山大猩猩、红毛猩猩以及几种长臂猿,只能在动物园看到了。要是我们以第三种黑猩猩的立场,决定救助另外两种黑猩猩,那么富裕国家的同胞,必须挑起主要的财务担子。从猿的观点来看,我们最近才搞清楚的“三种黑猩猩的故事”,其发挥的最重要功能,是决定我们面对那笔预算的态度。
然后再问自己:为什么那些猿被关在笼里让人参观?为什么还有些猿被用来做医学实验?可是对人,就不容许那样对待。假定学术界有一天发现:猿的基因组中,有99.9%的基因与人类的完全相同,那微小的0.1%,决定了人与猿之间的重大差异。那么你还会同意把猿关在笼子里供人参观,或者拉去做医学实验吗?再想想我们那些心智失常的不幸同胞,他们解决问题的能力很低,甚至无法照顾自己、无法沟通与发展社会关系、对疼痛的感觉也比不上猿。可是我们无法拉他们去做医学实验;猿就可以。其中的逻辑是什么?
[1] 盖伦(Galen),公元129—200年,约与华佗同时。——译者
下一回你逛动物园,请记得到各种猿类的笼子前走走。请想象那些猿身上的毛都脱落了,再想象它们附近另有一个笼子,其中关了几个咱们的弟兄,他们很不幸,给剥光了衣服,也说不出话来。现在请猜猜看,那些猿在遗传上与人究竟有多大的差别?猿与人的基因组中,有多少基因是共有的呢?占基因组的百分之几?10%?50%?还是99%?